細(xì)胞骨架(cytoskeleton)是指真核細(xì)胞中的蛋白纖維網(wǎng)絡(luò)結(jié)構(gòu)。發(fā)現(xiàn)較晚��,主要是因為一般電鏡制樣采用低溫(0-4℃)固定��,而細(xì)胞骨架會在低溫下解聚�����。直到20世紀(jì)60年代后����,采用戊二醛常溫固定,才逐漸認(rèn)識到細(xì)胞骨架的客觀存在���。
細(xì)胞骨架不僅在維持細(xì)胞形態(tài)����,承受外力���、保持細(xì)胞內(nèi)部結(jié)構(gòu)的有序性方面起重要作用�,而且還參與許多重要的生命活動(圖1),如:在細(xì)胞分裂中細(xì)胞骨架牽引染色體分離����,在細(xì)胞物質(zhì)運(yùn)輸中,各類小泡和細(xì)胞器可沿著細(xì)胞骨架定向轉(zhuǎn)運(yùn)�����;在肌肉細(xì)胞中�,細(xì)胞骨架和它的結(jié)合蛋白組成動力系統(tǒng);在白細(xì)胞的遷移�����、精子的游動�、神經(jīng)細(xì)胞軸突和樹突的伸展等方面都與細(xì)胞骨架有關(guān)。另外�,在植物細(xì)胞中細(xì)胞骨架指導(dǎo)細(xì)胞壁的合成。
細(xì)胞骨架由微絲(microfilament)���、微管(microtubule)和中間纖維(intemediate filament)構(gòu)成�。微絲確定細(xì)胞表面特征��,使細(xì)胞能夠運(yùn)動和收縮�。微管確定膜性細(xì)胞器(membrane-enclosed organelle)的位置和作為膜泡運(yùn)輸?shù)膶?dǎo)軌���。中間纖維使細(xì)胞具有張力和抗剪切力。
微絲�����、微管和中間纖維位于細(xì)胞質(zhì)中���,又稱胞質(zhì)骨架,它們均由單體蛋白以較弱的非共價鍵結(jié)合在一起�����,構(gòu)成纖維型多聚體���,很容易進(jìn)行組裝和去組裝�,這正是實現(xiàn)其功能所必需的特點����。
廣義的細(xì)胞骨架還包括核骨架(nucleoskeleton)、核纖層(nuclear lamina)和細(xì)胞外基質(zhì)(extracellular matrix)�����,形成貫穿于細(xì)胞核、細(xì)胞質(zhì)�、細(xì)胞外的一體化網(wǎng)絡(luò)結(jié)構(gòu)。
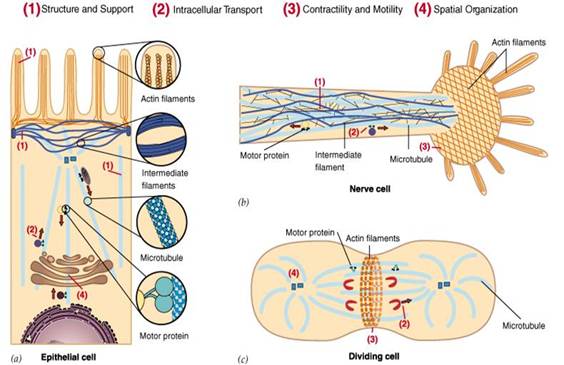
圖1為細(xì)胞骨架的主要功能
細(xì)胞骨架是細(xì)胞功能的結(jié)構(gòu)基礎(chǔ)����,在外界刺激或在細(xì)胞分裂、分化過程中��,細(xì)胞形態(tài)發(fā)生相應(yīng)的變化���,這和信號傳導(dǎo)調(diào)控細(xì)胞骨架的動態(tài)學(xué)變化分不開的��。細(xì)胞骨架作為真核細(xì)胞中的動態(tài)蛋白纖維網(wǎng)結(jié)構(gòu)���,由微絲(microfilaments, MF)、微管(microtubules, MT)和中間纖維構(gòu)成(intermediate filaments, IF)�。MF是由肌動蛋白聚合而成的多聚體, 廣泛存在于真核細(xì)胞應(yīng)力纖維、黏著斑��、偽足和收縮環(huán)中, 其結(jié)構(gòu)組成單元球狀肌動蛋白(globular actin, G-actin)在ATP和多種微絲相關(guān)蛋白的作用下, 通過與絲狀肌動蛋白(filamentous actin, F-actin)不斷組裝和解聚的轉(zhuǎn)換, 參與調(diào)控細(xì)胞形態(tài)改變��、黏附�、遷移和胞質(zhì)分裂等行為。MT由α-tubulin和β-tubulin聚合組成, 在微管相關(guān)蛋白的作用下組裝形成中空管狀結(jié)構(gòu), 在細(xì)胞內(nèi)物質(zhì)運(yùn)輸與胞質(zhì)分裂中扮演重要角色�。MF和MT細(xì)胞骨架相互聯(lián)系����、相互依賴, 共同參與調(diào)控細(xì)胞行為(Bailly M,2002��; Miranti C.K,2002)���。
細(xì)胞遷移是惡性腫瘤侵襲和轉(zhuǎn)移中的關(guān)鍵步驟����,細(xì)胞骨架和其結(jié)合蛋白是細(xì)胞遷移過程的物質(zhì)基礎(chǔ)�,細(xì)胞定向運(yùn)動需要細(xì)胞骨架的參與��,尤其是由肌動蛋白組成的微絲骨架�。MF和MT不僅在穩(wěn)定細(xì)胞形態(tài)、承受外力刺激�、維持細(xì)胞內(nèi)部結(jié)構(gòu)有序性中起重要作用, 而且還參與調(diào)控細(xì)胞遷移、黏附�����、分裂以及細(xì)胞內(nèi)信號傳遞����。其中, cAMP介導(dǎo)的信號通路與細(xì)胞骨架之間存在的互動效應(yīng)與細(xì)胞增殖����、凋亡�����、黏附����、遷移等細(xì)胞行為密切相關(guān), cAMP依賴的蛋白激酶A(PKA)作為cAMP依賴的主要靶向蛋白, 參與了這些過程。PKA通過磷酸化作用激活RhoA, RhoA激活后除了進(jìn)一步激活下游通路外, 還增強(qiáng)與Rho二磷酸鳥苷解離抑制因子(RhoGDI) 的結(jié)合能力, 從而反向抑制RhoA活性(Mehlen, P�,2006)。
整合素是細(xì)胞表面一類重要的兼具粘附和信號轉(zhuǎn)導(dǎo)功能的受體�,它是由α和β亞基通過非共價鍵組成的異二聚體。細(xì)胞外基質(zhì)蛋白���,如纖粘連蛋白���、層粘連蛋白和膠原等是其主要配體;其次還有一些細(xì)胞表面分子����,可溶性蛋白等也可與整合素結(jié)合。通過胞外段與胞內(nèi)段與細(xì)胞骨架、信號轉(zhuǎn)導(dǎo)分子和其它一些蛋白相結(jié)合�,整合素象是一座橋梁介導(dǎo)了細(xì)胞內(nèi)外之間的雙向信息傳遞。整合素可以與細(xì)胞骨架之間的雙向信號轉(zhuǎn)導(dǎo)�,(一)從細(xì)胞骨架到整合素,整合素在沒有與結(jié)合時是分散在細(xì)胞表面的����,而且不與肌動蛋白細(xì)胞骨架相連,一旦與ECM中的配體結(jié)合�,則通過其胞內(nèi)域與細(xì)胞骨架發(fā)生聯(lián)系,并發(fā)生簇集��。(二)從整合素到細(xì)胞骨架,整合素可反饋調(diào)節(jié)細(xì)胞骨架的組裝�����,整合素介導(dǎo)的粘附自身就可以激活Rho家族小G蛋白(Mitra S.K���,2005;Larsen M,2003����; Turner C.E,2000)。